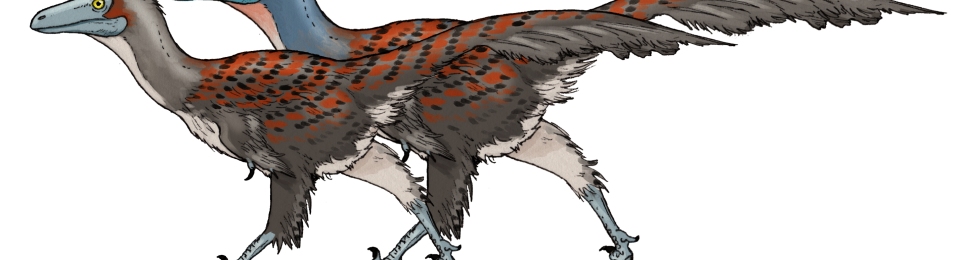

Mother Tianyulong is not happy. But what are each of her colour patterns for?
In the first two parts of this series, we went through different pigments that make up the rainbow of living bird colour, and discussed which of them were probably present on the skins and feathers of non-extinct dinosaurs.
In the end, we concluded that almost any colour, from black to white and orange to green, is possible. However, we paid no attention to what kind of colours are ecologically plausible. This article goes through different evolutionary pressures that shape animal colour – camouflage, impressing mates, keeping feathers intact.
Whole books and life’s works have been spent on the subject, so this article will necessarily be only a brief introduction. I will mostly use dinosaurs (both non-avian and living) as examples, but much of this also applies to other groups of extinct animals.
Something to hide
Crypsis is any ability of an animal to avoid detection by other animals. If there is no reason to stand out, most animals should be expected to blend to their surroundings reasonably well – the better the greater the need to hide. For example, large terrestrial predators such as big cats are mostly well camouflaged to be able to stalk their prey unseen, while the same is vital to ground-nesting birds for the opposite reason.
One of the ways to achieve this is countershading. The phenomenom is sometimes called Thayer’s law after the first person to document it in detail, the British artist Abbott Handerson Thayer in 1909 (so who says artists are not needed in science?). Animals tend to be lighter on the underside to compensate for the shadow cast by their own body. This colour difference makes it harder for others to pinpoint their shape.

Tsintaosaurus illustrates how crypsis can vary between vegetation types.
Generally pigmentation varies with latitude. Close to the equator, animals tend to be darker in colour, states Gloger’s rule. Oddly, this rule applies to both birds and mammals, but for different reasons! Hairless mammals, such as humans, are darker close to the equator to shield themselves from ultraviolet radiation (Chaplin 2004). This might also be true for scaly dinosaurs, though their scales might protect them from excessive radiation regardless of their colour. Birds have darker feathers because black eumelanin protects them from bacterial degradation in humid environments, such as equatorial rainforests (Burtt & Ichida 2004). In hairy mammals, this tendency probably has something to do with dark hues simply being a better camouflage under a tropical forest canopy (Kamilar & Bradley 2011). It’s by no means a law: there obviously are plenty of dark-pigmented animals – say, musk oxen or ravens – in high latitudes and open environments, and white birds in the tropics.

A good example of the power of countershading. The chital deer (Axis axis) on the background are effectively camouflaged because their white bellies counteract the shadow cast by their body. In the deer standing up, the white belly is glaringly visible. The white spotting is distractive colouration. Photo: Yathin S Krishnappa / Wikipedia. CC 3.0 licence.
Making a good impression
Sexual selection occurs when animals either actively choose with whom to mate, or compete with one another from the opportunity. It’s usually the explanation for extravagant colour and seemingly unnecessary beauty in nature. The male peacock is a classic example, but colours produced by sexual selection can also be subtle and are not restricted to males.
Sexual dimorphism, or differences between males and females, often evolve when the sexes have a different level of parental investment – one of the sexes does most of the work. The sex doing most of the work has more interest in being choosy, as it has more to lose from picking a mate with bad genes. Most often, the hard-working parent is the female for the sheer necessity of her producing large, expensive eggs compared to tiny sperm (which is called Bateman’s principle. There are a lot of old men immortalized in ecological terminology).
The colours and ornaments produced by sexual selection do not need to make any sense. In fact, being cumbersome, expensive and dangerous is the very reason they evolve. These acts as honest signals of the animal’s condition and good genes: a weak or sickly individual could not produce the ornaments, or would quickly get eaten because of them. The animals themselves, obviously, do not think about the genetic quality of their future offspring. As far as anyone knows, they simply have a sense of beauty.

Mandarin ducks form monogamous pairs, but the male takes no part in brooding and leaves before ducklings even hatch. This mismatch of workload is probably why such a large dimorphism has evolved. Photo: Francis C. Franklin / Wikimedia Commons. CC 3.0 licence.
With bigger differences of parental investment, higher level of promiscuity and/or tougher competition between males, a more pronounced dimorphism tends to evolve. It is most obvious in lekking birds, such as many ducks, landfowl (including peacocks) and bustards. The selection for more extravagant ornaments will stop when natural selection opposing them grows equally strong. That is, when the most beautiful male becomes more likely to die because of its ornaments than get to mate because of them. This spot is different for each species, depending on the subtleties of their habits, social life and predation pressure.
In species with single parent males, the roles are sometimes reversed. When males do most of the work related to breeding, it is the females who compete for their attention, and may evolve to be larger and more colourful. One such species is the red-necked pharalope (Phalaropus lobatus), a small Arctic wader (Hildén & Vuolanto 1972). This kind of arrangements could certainly have occurred in some dinosaur species. On the other hand, they are all but impossible in mammals, where the female is tied to her young by lactation.

A few approaches to sexual dimorphism, as illustrated by Stenonychosaurus. “Socially monogamous” refers to animals that raise young together as pairs, but have plenty of affairs on the side (such as many small songbirds). Dimorphism like this also occurs in monogamous birds with clearly different roles in nesting (such as sparrowhawks).
Some dinosaurs evolved equal
In monogamous bird species, dimorphism is generally lower, but it doesn’t mean sexual selection does not happen – it’s just mutual. When both parents work hard to raise young, they both have reason to be picky – and accordingly, reason to be pretty. There is also the whole range between these two extremes, that can and should also be explored through paleoart.
In many monogamous birds, such as most parrots, penguins, starlings and hornbills, both sexes are beautifully coloured, but the differences between them can be very subtle: in budgerigars, the only visible difference is colour of the cere, or the skin area containing the nostrils. Sometimes, the differences can only seen in the ultraviolet range. These birds can be colourful, because neither parent needs to expose itself to danger while brooding eggs: penguins live in areas without land predators, and the others nest in holes. The need for crypsis limits colouration in animals that need to brood out in the open, which includes at least oviraptorosaurs.
In flocking or herding animals, a lack of dimorphism can also be explained by the need to blend together. Individuals that stand out are often targeted by predators. This might have been true for some dinosaurs or pterosaurs.
One possible explanation for the lack of confirmed dimorphism in non-avian dinosaurs might actually be that many of them were more of less monogamous, or that mutual sexual selection was otherwise predominant in them (Hone et al. 2011). Monogamy is the most common social system in living birds, after all.
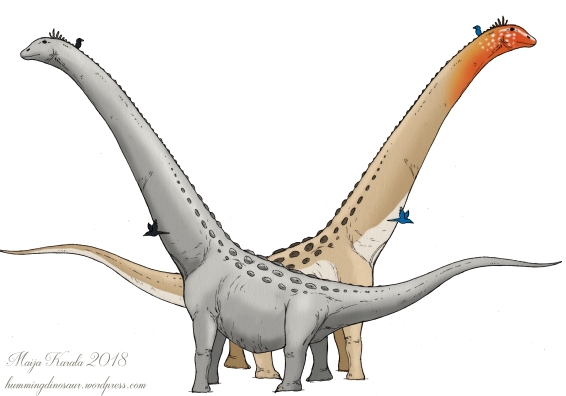
Two ways to go with gigantic dinosaurs.
Elephant grey or bright as a parrot?
Today, most large mammals are relatively drab in colour. Their sexual ornaments are in the form of horns, antlers, tusks, or manes, not bright pigments. This is often also followed in paleoart depicting large dinosaurs, given that mammalian megafauna is their closest living ecological equivalent. Sauropods with the grey and wrinkly skin of an elephant are a persistent paleoart trope.
However, there is a problem: all mammals are partially colour-blind compared to birds, and very likely other dinosaurs. So which is more important in determining the colouration of big mammals: ecology or vision? Are living birds only colourful because they can afford it, being flighted and relatively safe from predators?
Large flightless birds may offer a hint, given that they ARE in fact megafaunal dinosaurs. Of the four genera alive, one (the rhea) is completely drab. Emus have pale blue heads, male ostriches have showy black-and-white plumage with a pink or blue head, and cassowaries are extremely colourful. The full scale from drab to rainbow seems to be present even in this handful of species.
Another line of thought is to look at other living flightless diapsids. Can reptiles afford to be colourful, despite being tied to the ground and vulnerable to predators? Some apparently do, given the existence of many colourful agamas, chameleons, skinks, and geckos.
In conclusion, while some non-avian dinosaurs and other extinct diapsids likely were mostly grey or brown, there is no real reason to think there weren’t also showy ornaments and colourful species.
Warning!
Aposematic colour is most well known in small animals like wasps, but it also exists in birds and mammals, and could have been present in some non-avian dinosaurs. It serves to warn a potential predator that the animal is not helpless.
For the orange-and-black New Guinean pitohuis, it is because they are actually poisonous. They carry similar toxins as poison dart frogs, produced by the insects they eat (Dumbacher et al. 1992). These passerine birds are reportedly toxic enough that the hands of the ornithologists handling them went numb.
In mammals, aposematism may be a warning for being spiny (such as in the streaked tenrecs), for being smelly (in skunks), or even for having a really bad temper (in the honey badger). While smelly anal glands are unique to mammals, I’m fairly sure some dinosaurs could have protected themselves by being spiny or nasty, and had the colour to match.
Now let’s get back to the Tianyulong piece that started this article. Here is the reasoning behind each colour and pattern choice:
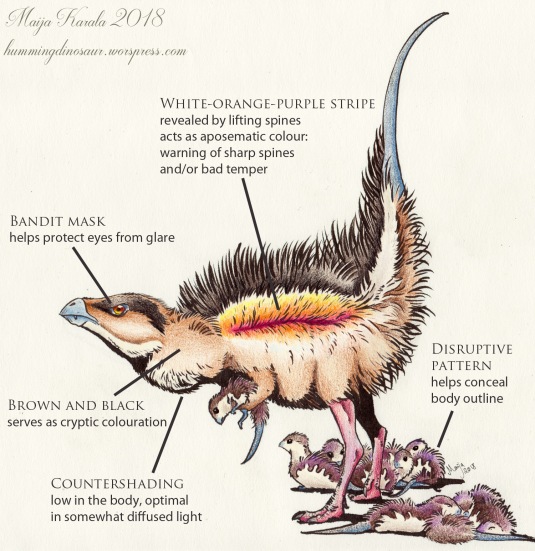
Think of the zebras! Everything I have described here is just general rules of thumb. While most living species of bird and mammal can be said to roughly follow them, evolution has produced plenty of subtle variations and quirky solutions that have left biologists scratching their heads for decades, for example the true purpose of zebra stripes. So don’t be afraid to use some creativity here and there.
References:
The general principles of crypsis, sexual selection, and aposematic colour can be found in many biology and behavioural ecology textbooks.
Burtt & Ichida 2004: Gloger’s rule, feather-degrading bacteria, and color variation among song sparrows. The Condor: Ornithological Applications.
Chaplin 2004: Geographic distribution of environmental factors influencing human skin coloration. American Journal of Physical Anthropology.
Dumbacher et al. 1992: Homobatrachotoxin in the Genus Pitohui: Chemical Defense in Birds? Science.
Hildén & Vuolanto 1972: Breeding biology of the Red-necked Phalarope Phalaropus lobatus in Finland. Ornis Fennica.
Hone et al. 2011: Does mutual sexual selection explain the evolution of head crests in pterosaurs and dinosaurs? Lethaia.
(A highly interesting Tetrapod Zoology article in the paper and its subject by Darren Naish can be read here)
Kamilar & Bradley 2011: Interspecific variation in primate coat colour supports Gloger’s rule. Journal of Biogeography.
Wikipedia: Crypsis, Countershading, Sexual selection, Aposematism, Gloger’s rule.

.jpg){kind=link}
{kind=link}
I love this!
I _cannot_ thank you enough. Thank you!!
Just what I’ve been looking for. Great information for a book I’m working on right now.
Hello Maija. I like your work. I have been looking for an illustrator for a children’s picture book for some time. Do you ever draw humans? If so, and if you think you might be interested in this project, please message abinns@uclan.ac.uk
Zoology is the study of animals. Zoologist studied animals, their behavior & interactions with ecosystems. Read articles of zoology on https://everydayscience.blog/category/zoology/
Ciao , penso che i saw che avete visitato il mio sito quindi i
got qui a ritorno il favore sono?. tentare di ricerca cose a
migliorare mia sito presumo ! suo ok di fare uso di alcuni
del vostre idee !! Maramures Grazie, buona giornata!